
ID
64
Cancer Name
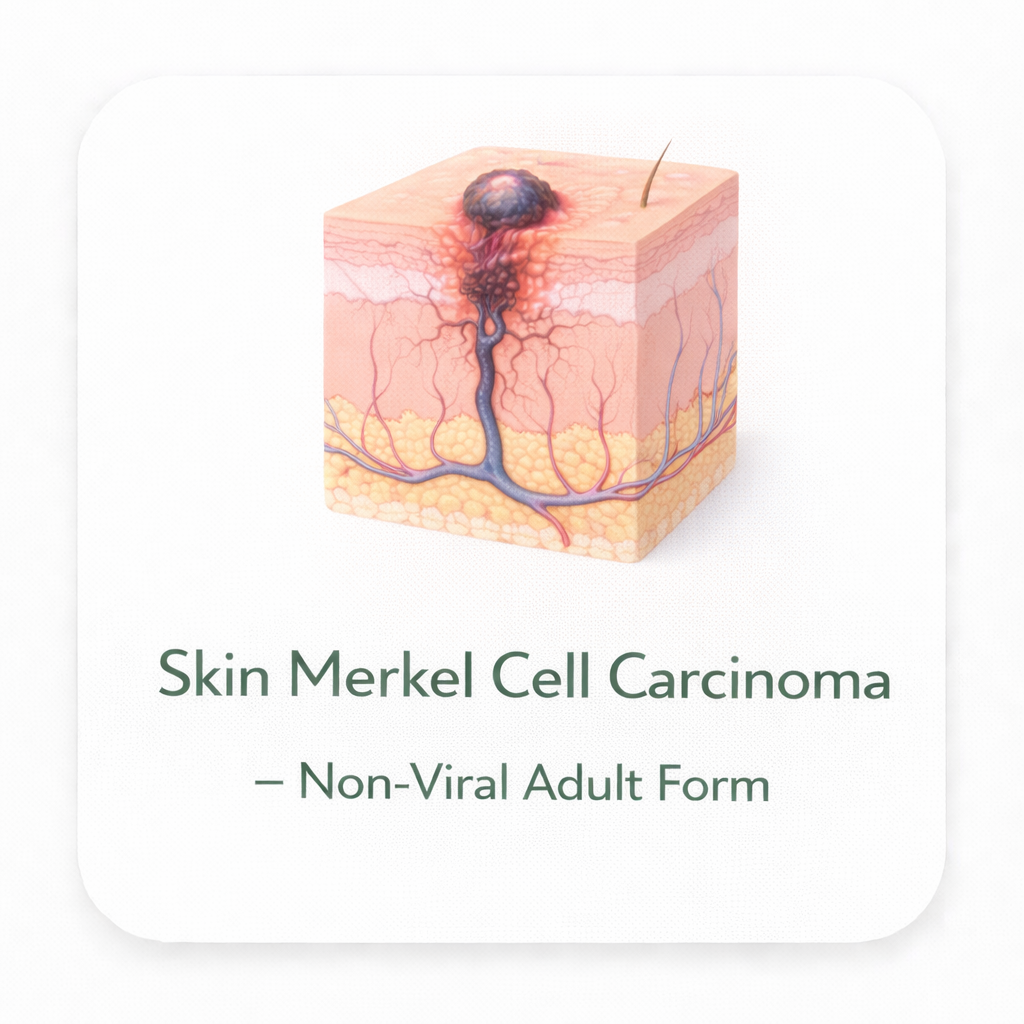
Skin Merkel Cell Carcinoma – Non-Viral Adult Form
Main Grouping
Integumentary
Organ System
Skin
Cell Origin
Neuroendocrine cells
Pathways Affected
Non-viral Merkel cell carcinoma involves a pathway landscape dominated by UV-induced TP53/RB1 dual inactivation, PI3K/AKT/mTOR pathway activation (~53% of cases), Wnt/beta-catenin signaling, NOTCH pathway suppression, DNA repair pathway deficiency, NF-kB survival signaling, and the PD-1/PD-L1 immune evasion checkpoint.
The TP53/RB1/cell cycle checkpoint pathway is the defining dual tumor suppressor inactivation in non-viral UV-induced MCC: UV radiation creates C>T transitions at dipyrimidine sites (the UV mutational signature) — in non-viral MCC, TP53 mutations occur in approximately 71-80 percent of cases eliminating the p53 DNA damage response checkpoint that would normally arrest UV-damaged cells for repair or trigger their apoptosis; UV-induced TP53 mutations are predominantly C>T transitions at TpC or CpC sites; RB1 mutations/deletions occur in approximately 71 percent of non-viral MCC cases — loss of RB1 protein removes the only brake on E2F transcription factors creating constitutive cell cycle entry through CDK4/6-cyclin D1 driven E2F activation; concurrent TP53 and RB1 loss creates a cell that cannot arrest at either the G1/S checkpoint (RB1 lost) or activate apoptosis in response to DNA damage (TP53 mutated) — the dual inactivation pattern seen in the most aggressive non-viral MCC; curcumin activates p53 pathway signaling in cancer cell models through MDM2 inhibition, promoting p53-independent apoptosis through Bax/BCL2 ratio modulation; quercetin inhibits RB1/E2F-driven cell cycle progression in cancer cell models.
The PI3K/AKT/mTOR pathway is activated in approximately 53 percent of non-viral MCC through multiple convergent genetic mechanisms confirmed by comprehensive genomic profiling of MCC (PMC5029639): PIK3CA activating mutations (~10-15% of non-viral MCC); PTEN loss/mutations (~15-20%) — PTEN is a phosphatase that degrades PIP3 suppressing AKT activation, its loss creates constitutive AKT/mTOR signaling; AKT2 amplification (~subset); FBXW7 mutations disrupting mTOR negative regulation (~subset); constitutive mTORC1 activation from PI3K/AKT/mTOR drives ribosome biogenesis, protein synthesis, and the large nucleoli characteristic of MCC morphology; PI3K pathway activation was confirmed in human MCC cell lines and tumors with a complete clinical response reported in a stage IV MCC patient treated with PI3K inhibitor (PMC7264292); quercetin inhibits PI3K/AKT/mTOR in skin cancer cell models; curcumin inhibits PI3K/AKT and mTOR through AMPK pathway activation in cancer cell models targeting the dominant ~53% PI3K/AKT/mTOR pathway in non-viral MCC.
The Wnt/beta-catenin pathway is active in MCC cells and skin cancers: beta-catenin is expressed in MCC cells; DVL2/beta-catenin/cyclin D1/Cox2 Wnt target genes drive MCC cell cycle progression and survival; cyclin D1 (CCND1) is a CDK4/6 activator that phosphorylates RB1 — in non-viral MCC where RB1 is already mutated, cyclin D1 upregulation by Wnt target gene activation further amplifies E2F-driven transcription; curcumin and quercetin synergistically downregulated DVL2, beta-catenin, cyclin D1, Cox2, and Axin2 in A375 skin cancer cells confirmed by Western blot (PubMed30599890) — directly targeting the Wnt/beta-catenin/cyclin D1 axis relevant to MCC cell proliferation; curcumin inhibits Wnt/beta-catenin through DVL2 downregulation and beta-catenin reduction in skin cancer cell models.
The NOTCH pathway is suppressed rather than activated in non-viral MCC — NOTCH1/2 inactivating mutations create loss of the tumor-suppressive NOTCH signaling in MCC: normal NOTCH1 signaling in skin keratinocytes promotes differentiation and growth arrest; NOTCH1 inactivating mutations in non-viral MCC (identified in comprehensive genomic profiling PMC5029639) remove this differentiation/growth arrest signal creating a stem-cell-like undifferentiated proliferative state; this is the opposite of Notch pathway activation seen in many other cancers; the NF-kB pathway is constitutively active in MCC cells creating survival and anti-apoptotic gene expression including Bcl-2, Bcl-XL, Survivin; curcumin inhibits NF-kB through IKK-beta suppression targeting the constitutive NF-kB survival program in MCC.
Description
Merkel cell carcinoma (MCC) is an uncommon but highly aggressive primary neuroendocrine carcinoma of the skin. It is the second most lethal skin cancer after melanoma with an overall 5-year mortality rate of approximately 33 to 46 percent for all stages. In the United States, approximately 3,000 new cases of MCC are diagnosed annually as of the most recent available data, with incidence increasing approximately 3 to 5 percent annually since 1986 correlating with the aging and immunosuppressed population as well as cumulative UV exposure. Globally, MCC accounts for approximately 5,000 to 7,000 new cases annually. Non-viral (MCPyV-negative) MCC accounts for approximately 20 to 40 percent of all MCC and is the predominant form in elderly adults over age 70 with extensive lifetime sun exposure. Non-viral MCC carries a 5-year overall survival of approximately 25 to 40 percent — approximately 3-fold worse than viral-positive MCC which has a 5-year OS of approximately 45 to 65 percent.
The demographic profile of non-viral adult MCC: predominantly affects older adults (median age approximately 75-80 years in non-viral form); male-to-female ratio of approximately 2:1; strong association with cumulative UV radiation exposure and sun-damaged skin (actinic keratoses, solar elastosis in adjacent skin); geographic high-incidence in Australia, New Zealand, and UV-intense regions; risk factors include age >65, immunosuppression (organ transplant recipients have approximately 10-fold increased risk), chronic UV exposure, and fair-skin phototypes I-II.
The staging of MCC follows the American Joint Committee on Cancer (AJCC) 8th edition staging: stage I (primary ≤2 cm, node negative) 5-year OS approximately 55-65 percent; stage II (primary >2 cm, node negative) 5-year OS approximately 45-55 percent; stage III (regional node positive) 5-year OS approximately 20-35 percent; stage IV (distant metastasis) 5-year OS approximately 0-18 percent.
The molecular biology of non-viral MCC is defined by the UV damage mutational signature creating approximately 1,121 somatic single nucleotide variants per exome — dual TP53/RB1 inactivation in approximately 71 percent of cases, PI3K/AKT/mTOR pathway activation in approximately 53 percent, NOTCH pathway suppression, and chromatin modification gene mutations; this UV-induced mutation landscape contrasts fundamentally with viral MCC where viral T antigens inactivate RB1 and p53 without UV mutagenesis.
Published laboratory research documents curcumin and quercetin synergistically inhibiting cancer cell proliferation in A375 human skin cancer cells — inhibiting Wnt/beta-catenin signaling pathway proteins (DVL2, beta-catenin, cyclin D1, Cox2, Axin2) and inducing apoptosis through BCL2 downregulation and caspase 3/7 activation via PARP cleavage confirmed by MTT assay, colony proliferation assay, and Western blot (PubMed30599890) — targeting the Wnt/beta-catenin pathway active in skin cancer cells and the BCL2-dependent apoptosis resistance documented in MCC, and the cyclin D1-driven cell cycle progression relevant to RB1-loss-driven cell cycle deregulation in non-viral MCC.
Plant-Based Description
Whole-food plant-based dietary patterns provide phytochemicals with documented activity in skin cancer cell lines directly applicable to non-viral Merkel cell carcinoma through its dominant PI3K/AKT/mTOR and Wnt/beta-catenin pathways, BCL2-dependent apoptosis resistance, and UV-induced TP53/RB1 loss. Curcumin and quercetin synergistically inhibited cancer cell proliferation in A375 human skin cancer cells through Wnt/beta-catenin signaling pathway modulation (DVL2, beta-catenin, cyclin D1, Cox2, Axin2 downregulated) and apoptosis induction through BCL2 downregulation and caspase 3/7 activation via PARP cleavage confirmed by MTT assay, colony proliferation assay, and Western blot (PubMed30599890) — directly targeting the Wnt/cyclin D1 cell cycle axis relevant to RB1-loss-driven MCC proliferation and BCL2-dependent apoptosis resistance; curcumin additionally inhibits PI3K/AKT, mTOR, NF-kB, and MAPK/ERK in skin cancer cell models targeting the ~53% PI3K/AKT/mTOR pathway frequency in non-viral MCC; EGCG from green tea inhibits PI3K/mTOR, NF-kB, and activates Nrf2/ARE in skin cancer cell models; sulforaphane from cruciferous vegetables activates Nrf2/ARE and provides UV-induced DNA damage repair support; resveratrol inhibits Wnt/beta-catenin, NF-kB, and PI3K/AKT in skin cancer cell models.
Plant Chemistry Detail
Curcumin and quercetin have confirmed anti-skin-cancer cell line activity in a published study (PubMed30599890) using A375 human melanoma skin cancer cells — sharing UV-induced oncogenesis pathways with non-viral MCC. In this confirmed study: curcumin inhibited proliferation in A549 (lung) and HCT116 (colon) cancer cell lines with IC50 of 3 to 8.5 μM confirmed by MTT assay; quercetin showed stronger inhibition of cell proliferation than berberine confirmed; combination of curcumin and quercetin in four cancer cell lines including A375 skin cancer cells showed synergistic effect on cell proliferation with several-fold decreases in IC50 confirmed by MTT and colony proliferation assays; investigation of the mechanism in A375 skin cancer cells confirmed that inhibition of cell proliferation occurred through downregulation of Wnt/beta-catenin signaling pathway proteins DVL2 (Dishevelled 2, the proximal Wnt signal transducer), beta-catenin, cyclin D1 (CDK4/6 activator and RB1 phosphorylation substrate), Cox2, and Axin2 confirmed by Western blot; both curcumin and quercetin induced apoptosis by downregulating BCL2 confirmed by Western blot and inducing caspase 3/7 through PARP cleavage confirmed — demonstrating that curcumin and quercetin target both the proliferative Wnt/beta-catenin/cyclin D1 axis and the anti-apoptotic BCL2 survival program in A375 human skin cancer cells; the mechanism is directly relevant to non-viral MCC because: (1) cyclin D1 downregulation addresses the E2F-driven proliferative program resulting from RB1 loss (71% of non-viral MCC); (2) BCL2 downregulation addresses BCL2 overexpression documented in MCC; (3) Wnt/beta-catenin modulation addresses the Wnt pathway active in MCC cells; (4) the UV-induced mutational signature of A375 melanoma parallels the UV-induced mutational landscape of non-viral MCC.
EGCG from green tea inhibits PI3K/mTOR and NF-kB in skin cancer cell models — directly targeting the ~53% PI3K/AKT/mTOR pathway frequency and NF-kB survival signaling in non-viral MCC; EGCG also activates Nrf2/ARE providing cytoprotection against UV-induced oxidative DNA damage in sun-exposed skin cells; EGCG inhibits STAT3 phosphorylation in skin cancer cell models targeting JAK/STAT3 pro-survival signaling. Sulforaphane from broccoli and cruciferous vegetables activates Nrf2/ARE in keratinocytes and skin cells — upregulating NQO1, HO-1, GPx, and glutathione biosynthesis enzymes that defend against UV-induced reactive oxygen species and UV-induced DNA damage; sulforaphane inhibits NF-kB in skin cancer cell models. Apigenin from parsley inhibits PI3K/AKT, STAT3, and cell cycle progression in skin cancer cell models targeting the PI3K/AKT pathway and STAT3 activation in MCC; apigenin activates p53 in TP53-wild-type cell models and induces p53-independent apoptosis in TP53-mutant cell models. Resveratrol inhibits Wnt/beta-catenin, NF-kB, PI3K/AKT, and MAPK/ERK in A375 and other skin cancer cell models. Luteolin inhibits PI3K/AKT and mTOR in skin cancer and neuroendocrine tumor cell models targeting the ~53% PI3K/AKT/mTOR pathway in non-viral MCC. Allicin from garlic induces apoptosis and inhibits NF-kB in skin cancer cell models.
Nutritional Focus
Nutritional focus in non-viral Merkel cell carcinoma research is led by curcumin from turmeric and quercetin from onions with confirmed anti-skin-cancer cell line activity in A375 human skin cancer cells: curcumin and quercetin synergistically inhibiting A375 skin cancer cell proliferation through Wnt/beta-catenin signaling pathway modulation — downregulating DVL2, beta-catenin, cyclin D1, Cox2, and Axin2 — and inducing apoptosis through BCL2 downregulation and caspase 3/7 activation via PARP cleavage confirmed by MTT assay, colony proliferation assay, and Western blot (PubMed30599890) — directly targeting the Wnt/cyclin D1 cell cycle axis compounding the E2F-driven proliferation from RB1 loss (~71% of non-viral MCC) and the BCL2-dependent apoptosis resistance documented in MCC; curcumin additionally inhibiting PI3K/AKT, mTOR, NF-kB, and Wnt/beta-catenin in skin cancer cell models — targeting the ~53% PI3K/AKT/mTOR pathway frequency in non-viral MCC; EGCG from green tea inhibiting PI3K/mTOR, NF-kB, STAT3, and activating Nrf2/ARE in skin cancer cell models — targeting both PI3K/mTOR and NF-kB survival signaling in MCC while activating antioxidant defense against UV-induced oxidative DNA damage; sulforaphane from cruciferous vegetables activating Nrf2/ARE and GCL/NQO1/HO-1 in keratinocytes targeting UV-induced ROS and oxidative DNA damage defense directly relevant to the UV-mutagenesis mechanism of non-viral MCC; apigenin from parsley inhibiting PI3K/AKT and STAT3 in skin cancer cell models; resveratrol inhibiting Wnt/beta-catenin, NF-kB, and PI3K/AKT in skin cancer cell models; quercetin and curcumin inhibiting IDO1 targeting tryptophan-kynurenine immunosuppression in the MCC tumor microenvironment; and dietary fiber from whole plant foods producing SCFAs that inhibit HDAC targeting the ASXL1/KMT2D chromatin modification gene mutations documented in non-viral UV-induced MCC.
Research Notes
Non-viral MCC epidemiology: ~20-40% of all MCC; ~3,000 total MCC cases/year US; incidence increasing ~3-5%/year; ~5,000-7,000 global cases/year; non-viral median age ~75-80 years; male:female ~2:1; strong UV exposure association; fair-skin phototypes I-II; immunosuppression risk 10-fold. 5-year OS all MCC ~54-64%; non-viral MCC ~25-40%; viral MCC ~45-65%; stage IV OS 0-18%. IHC: CK20+ (paranuclear dot), CK7-, Chromogranin A+, Synaptophysin+, CD56+, TTF-1-. Molecular non-viral MCC: UV mutational signature (C>T at dipyrimidine sites, SBS7a/SBS7b); median ~1,121 SSNVs/exome; TP53 mutations ~71-80% (UV-signature C>T transitions); RB1 mutations/deletions ~71%; concurrent TP53+RB1 loss characteristic; NOTCH1/2 inactivating mutations (high frequency); FAT1 mutations (high frequency); PI3K/AKT/mTOR pathway collectively ~53% (PIK3CA ~10-15%, PTEN ~15-20%, AKT2 amplification, FBXW7 mutations); CDKN2A/B loss; ATM/BRCA2/MSH2 DNA repair mutations ~29%; ASXL1/KMT2D chromatin gene mutations; MAP3K1/TRAF7 JNK pathway mutations; MYC amplification (subset); PD-L1 variable. PI3K pathway activation confirmed in MCC cell lines and tumors with complete clinical response to PI3K inhibitor in stage IV MCC (PMC7264292). Curcumin+Quercetin A375 skin cancer cells (PubMed30599890): MTT dose-dependent; colony proliferation confirmed; DVL2/beta-catenin/cyclin D1/Cox2/Axin2 downregulated Western blot; BCL2 downregulated; caspase 3/7 activated via PARP cleavage confirmed; synergistic effect confirmed.
Notes Visibility
show
Key Foods
Turmeric,Broccoli,Kale,Spinach,Brussels Sprouts,Cauliflower,Garlic,Yellow Onion,Carrot,Tomato,Beetroot,Cabbage,Blueberry,Pomegranate,Grape,Raspberry,Strawberry,Apple,Orange,Lemon,Soybeans,Edamame,Green Lentils,Black Beans,Chickpeas,Brown Rice,Quinoa,Oats,Wild Rice,Black Rice,Walnut,Almond,Brazil Nut,Flaxseed,Pumpkin Seeds,Chia Seeds,Sesame Seeds,Hemp Seeds,Shiitake,Maitake,Cremini,Portobello,Lions Mane,Green Tea,Ginger,Black Pepper,Garlic Powder,Parsley,Rosemary,Oregano,Sweet Potato
, Celery, Leek,Avocado,Artichoke,Radish,Tangerine, Red Onion
Linked Nutrients
vitamin-c,vitamin-e,vitamin-a,vitamin-b9,vitamin-b6,selenium,zinc,magnesium,calcium,potassium,iron,curcumin,quercetin,egcg,sulforaphane,apigenin,beta-carotene,lycopene,anthocyanins,beta-glucans,dietary-fiber
Last Updated
2025-10-13 10:19:09

